|
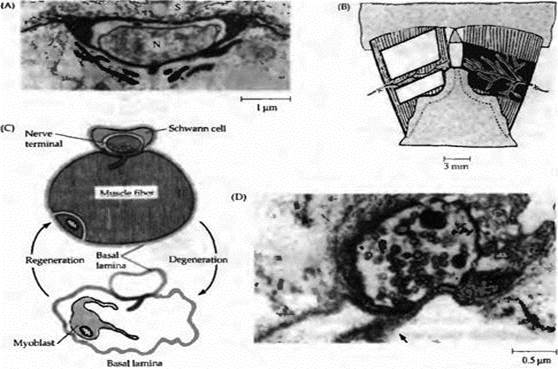
Для идентификации сигнала, связанного с базальной мембраной и инициирующего постсннаптическую дифференцировку, МакМахан и его коллеги использовали морского ската Torpedo califomica. Из электрических органов этого животного, родственных скелетной мышце, они приготовили экстракты, содержащие базальные мембраны. Добавленные к культуре мышечных волокон, экстракты симулировали эффекты синаптической базальной мембраны на регенерацию мышечных волокон, а именно индуцировали формирование кластеров АХ рецепторов вместе с другими компонентами постсинаптической мембраны (рис. 5).
Активный компонент экстрактов, названный агрином,
был очищен и охарактеризован, а у цыпленка, крысы и ската клонирована соответствующая кДНК. |
| |
Рис. 4. Базальная мембрана и регенерация синапсов. (А) Микрофотография нормального нервно-мышечного синапса лягушки, окрашенного рутением красным, показывающая базальную мембрану, погруженную в постсинаптические складки и окружающую шванновскую клетку (S) и нервное окончание (N). (В) Изображение кожно-грудной мышцы, показывающее замороженный (справа) или перерезанный (слева) участок, с целью вызвать локальное повреждение мышечных волокон. (С) Замораживание приводит к дегенерации и фагоцитозу всех клеточных элементов нервно-мышечного соединения, оставляя целой только базальную мембрану мышечного волокна и шванновской клетки. Новые нервно-мышечные синапсы создаются регенерирующими аксонами и мышечными волокнами. (D) Нерв и мышца были повреждены, регенерация мышечных волокон предупреждена рентгеновским облучением. В отсутствие мышечных волокон аксоны регенерировали, восстанавливали связь с исходными синаптическими зонами и формировали активные зоны. |
Результаты гибридизации in situ и иммуногистохимических исследований продемонстрировали, что агрин синтезируется мотонейронами, транспортируется по аксонам и, освобождаясь, индуцирует дифференцировку постсинаптического аппарата в развивающихся нервно-мышечных синапсах. Затем агрин становится частью синаптической базальной мембраны, где участвует в сохранении постсинаптического аппарата и запускает дифференцировку во время регенерации. |
Рис. 5. Аккумуляция AX рецепторов и ацетилхолинэстеразы в исходных синаптических зонах мышечных волокон, регенерирующих в отсутствие нерва. Мышца была заморожена, как на рис. 24.13В, но регенерация нерва была блокирована. Новые мышечные волокна сформировались в пределах оболочек базальной мембраны. (А и В) Ауторадиография регенерированной мышцы с окраской на холинэстераэу, для того чтобы выделить исходную синаптическую зону (в фокусе в части А), и помеченной радиоактивным -бунгаротоксином, чтобы определить положение АХ рецепторов (серебряные зерна в фокусе в части В). (С) Электронная микрофотография исходной синаптической зоны в регенерированной мышце, отмеченная пероксидазой хрена (HRP), конъюгированной с -бунгаротоксином. Распределение АХ рецепторов обнаруживается по высокой плотности продукта реакции с HRP, который позволяет различать поверхность мышечного волокна и синаптические складки. (D) Электронная микрофотография исходной синаптической зоны в регенерирующей мышце с окраской на холинэстеразу. |
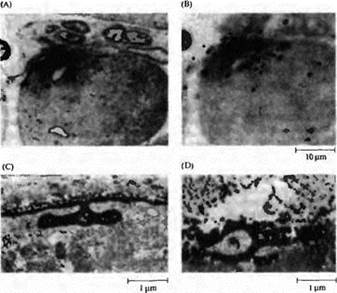
Исходная холинэстераза деградировала после заморозки мышцы. Таким образом, выявляемый продукт реакции обусловлен холинэстеразой, синтезированной и аккумулированной в исходной синаптической зоне регенерирующим мышечным волокном. |
|


