|
Для полного восстановления функции после повреждения нервной системы регенерирующие аксоны должны восстановить связи со своими прежними мишенями. Классические эксперименты Лэнгли, подтвержденные позже исследованиями одиночных клеток, показали, что регенерирующие преганглионарные аксоны нейронов автономной нервной системы реиннервируют соответствующие постганглионарные нейроны. В этом механизме имеет значение, по-видимому, сигнал о позиции нейрона, который оказывает влияние на формирование синапса между нейроном и клеткой-мишенью. |
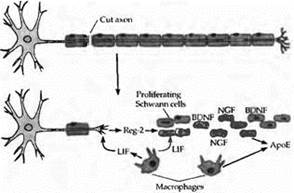
Рис. 2. Шванновские клетки способствуют регенерации аксонов в периферической нервной системе позвоночных. После аксотомии дистальный участок аксона и миелиновая оболочка дегенерируют и фагоцитируются. Пролиферация шванновских клеток стимулируется двумя цитокинами: фактором, ингибирующим лейкемию (LIF), из макрофагов и Reg-2 из окончаний аксонов. Экспрессия Reg-2 усиливается LIF. Пролиферирующие шваннов ские клетки синтезируют два нейротрофических фактора, BDNF и NGF, которые сохраняются на клеточной поверхности их низкоаффинными рецепторами. Нейротрофины поддерживают процесс регенерации аксонов и направляют их к соответствующим мишеням. Шванновские клетки и макрофаги синтезируют также аполипопротеин Ε (ΑροΕ), который способствует выживанию нейронов и регенерации их аксонов. |
|
Этот механизм справедлив как для симпатической, так и двигательной системы. Так, если межреберная мышца, извлеченная из передней части грудной клетки, трансплантируется в область шеи, то она преимущественно реиннервируется аксонами шейного симпатического ствола, расположенными в самых передних участках спинного мозга (рис. 3). Наоборот, мышцы, трансплантированные из задних областей тела, имеют тенденцию к реиннервации аксонами более дорзальньгх нейронов. Подобная тенденция установлена и для трансплантации симпатических ганглиев различного исходного расположения. У молодых крыс зависимость от позиции нейрона показана в реиннервации мышц, имеющих множественную иннервацию несколькими сегментами.
Нервно-мышечные синапсы у новорожденных крыс, головастиков и взрослых тритонов после поперечного пересечения двигательного нерва с высокой точностью воссоединяются с исходной синаптической зоной . Одним из механизмов избирательного восстановления синапсов является конкуренция между аксонами. В мышцах саламандры, иннервированных чужим аксоном, чужеродный синапс элиминируется после того, как нормальный нерв восстановил свой синаптический контакт. У взрослых млекопитающих чувствительные, двигательные и постганглионарные аксоны показывают более низкую избирательность по отношению к их исходным мишеням. Чужие нервы при иннервации мышечных волокон могут быть столь же эффективными, как и оригинальные нервные волокна. Более того, чужой нерв может даже вытеснить исходный аксон в интактных мышцах взрослой крысы). Избирательная регенерация у взрослых млекопитающих возможна, если периферический нерв был не перерезан, а передавлен так, чтобы сохранились эндоневрий и базальная мембрана шванновских клеток, окружавших интактные аксоны (см. рис. 4)). При таких условиях регенерирующие аксоны направляются вновь к своим исходным мишеням. Если эндоневрий был разрушен, например при перерезке нерва, регенерация происходит беспорядочно и аксоны часто создают синапсы с неподходящими мишенями. |
| |
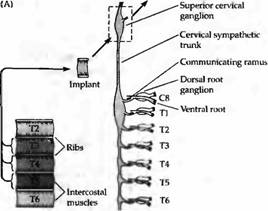
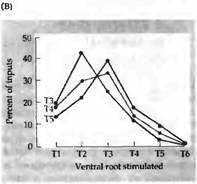
Рис. 3. Селективная реиннервация мышц, основанная на сегментарном происхождении. (А) Межреберная мышца из грудного сегмента ТЗ, Т4 или Т5 была трансплантирована на шею взрослой крысы и после удаления верхнего шейного ганглия была реиннервирована преганглионарными аксонами шейного симпатического ствола. Сегментарное происхождение входов было определено регистрацией активности мышц при стимуляции отдельных вентральных корешков, которые соединяют преганглиальные аксоны с нервным стволом. (В) Распределение входов к трансплантированным ТЗ, Т4 и Т5 мышцам. Передние мышцы склонны к реиннервации передними преганглиальными аксонами. |
|


