|
Стереотипно происходящее развитие сложного глаза дрозофилы представляет собой еще одну систему, в которой возможно прямое наблюдение для идентификации отдельных клеток и прослеживания их дальнейшего развития. Кроме того, генетика дрозофилы дает очень большие возможности для оценки роли происхождения клеток и индукционных взаимодействий между ними в дифференцировке нервной ткани. Возможно выделение мутантов, у которых нет определенного клеточного типа или у которых паттерн дифференцировки лишь незначительно нарушен, что позволяет определить их влияние на судьбу остальных клеток.
Взято тут http://mylnye-grezi.ru 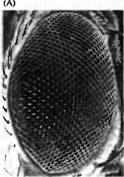
Рис. 4. Индукционные взаимодействия, регулирующие развитие фоторецепторных клеток у дрозофилы. (А) Сканограмма сложного глаза дрозофилы. Каждая фасетка представляет собой один омматидий. (В) Нормальное развитие дифференцировки восьми фоторецепторов в каждом омматидий. Seven/ess (sev-- ) и bride of seven (boss ) мутации нарушают дифференцировку R7. (С) Сигнальные каскады, регулирующие дифференцировку R7. Продукт boss гена, интегральный мембранный белок, экслрессируемый в R8 (Boss), активирует продукт гена sev, рецептора тирозинкиназы (SevRTK). Sev киназа запускает внутриклеточный сигнальный каскад, активирующий MAP киназу, которая имеет несколько мишеней. MAP киназа фосфорилирует протеин Van (который бы иначе блокировал дифференцировку), приводя к его распаду. MAP киназа также вызывает экспрессию белка Phyl который вместе со вторым протеином, Sina, приводит к распаду фактора транскрипции Ttk88 Ttk88 предотвращает нервную дифференцировку. MAP киназа также активирует Pntp2 и АР-1, два фактора транскрипции, которые способствуют нейрональной дифференцировке.
Подобная техника была использована в экспериментах Бензера, Реди, Рубина, Толимсона, Цирупски и их коллег, которые исследовали дифференцировку нейронов и сопутствующих (опорных) клеток глаза. Глаз дрозофилы состоит из кристаллоподобного набора повторяющихся единиц, называемых омматидиями (рис. 4A), каждый из которых содержит 8 фоторецепторов (R1-R8). Первая клетка, которая определяет начало дифференцировки в каждом омматидии, является одним из фоторецепторов, R8. Клетки R8 появляются хаотично в области нейроэпителия. Как только началась дифференцировка клетки R8, это приводит к ингибированию дифференцировки ее соседей в клетки R8. Затем дифферениировке подвергаются клетки R2 и R5, после чего R4, R1 и R6 и, наконец, R7 (рис. 4В). Были обнаружены две мутантные линии, в которых глаза развиваются нормально, за исключением того, что не образуется R7 (рис. 4В). Такие линии были названы sevenless (sev--) и brideof-sevenless (boss-- ) для обозначение отсутствия образования R7. При детальном исследовании этих мутантных линий впервые были изучены молекулярные механизмы того, как индукционные взаимодействия между клетками могут влиять на дальнейшее развитие клеток. Ген sevenless кодирует рецептор (называемый Sevenless, или Sev), для которого продукт экспрессии bride-of-sevenless гена, названный Boss, является лигандом. Во время развития R8 индуцирует образование R7. Это происходит, когда Boss, экспрессируемый на поверхности клетки R8, связывается с Sev, который расположен на клетке-предшественнице R7 (рис. 4С). Взаимодействие между Boss и Sev активирует внутриклеточный домен Sev рецептора, тирозинкиназу, которая инициирует сигнальный каскад в клетке R7, приводящий к ее дифференцировке. Сигнальный каскад является сложным путем, который включает в себя последовательную активацию серии протеинкиназ (ферментов, которые фосфорилируют белки), что приводит к ингибированию негативного влияния, а также увеличению позитивного влияния на экспрессию генов R7. Большое количество сигналов, которые приводят к изменениям в эспрессии генов, осуществляют это через тирозинкиназы, которые управляют данными клеточными каскадами.
|


