|
Происхождение клеток наиболее хорошо исследовано на примере простых беспозвоночных, таких как пиявка, кузнечик, плодовая муха и маленькая нематода Caenorhabditis elegans. В этих препаратах возможно проследить развитие каждой клетки и исследовать формирование таких характеристик, как свойства мембраны, трансмиттеры, рост аксонов и их ветвление. У С. elegans, которая содержит всего около 300 нейронов, эмбрион настолько маленький и прозрачный, что можно идентифицировать каждый нейрон и проследить его работу при помощи микроскопа. Альтернативным подходом может быть маркирование отдельных клеток и определение того, какие типы клеток из них получаются. Такого рода анализ, впервые предложенный Вайсблатом, Стентом и их коллегами для эмбрионов пиявки, включает в себя введение внутриклеточных маркеров, таких как флуоресцентный декстран или фермент пероксидаза хрена (HPR), в отдельные клетки и таким образом исследование дальнейшего потомства либо на живом эмбрионе, либо после окраски эмбриона, когда можно увидеть клетки, в которых находится фермент. Можно провести сравнительные эксперименты, вводя комплементарные цепи ДНК, кодирующие гены флуоресцентного белка, или создавая трансгенных животных, которые экспрессируют этот протеин.
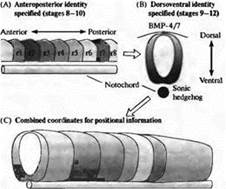
Рис. 4. Система координат пространственной информации в заднем мозге позвоночных, устанавливаемая в два этапа. (А) Сначала определяется рострокаудальная позиция, например по экспрессии гена Hox. (В) После этого дорзовентральная позиция определяется градиентами сигналов средней линии, такими как Sonic Hedgehog и BMP 4/7. (С) Результирующая двумерная система координат пространственной информации ограничивает возможный репертуар клеточной дифференцировки плюрипотентных клеток-предшественников.
Эксперименты подобного рода показывают, что у простых беспозвоночных вполне возможно воспроизвести определенную последовательность в делении и дифференцировке клеток. Таким образом, используя луч лазера можно вызывать гибель отдельных идентифицированных клеток с целью проследить, каким образом это повлияет на судьбу оставшихся клеток. В большинстве случаев выжившие клетки игнорируют потерю своего соседа, направление их развития определено автономно, на основании того, к какой клеточной линии они принадлежат. У подобных клеток экспрессия генов определяется факторами, которые изначально находятся в их цитоплазме или ядре и представляют собой независимые внутриклеточные сигналы. В других случаях, однако, потеря соседа может повлиять на судьбу выживших клеток. Таким образом, даже у животных, у которых имеются жестко определенные паттерны деления клеток, их дальнейшее развитие может изменяться в зависимости от индукционных взаимодействий.
|


