|
Для того чтобы вычислить точное значение мембранного потенциала для модели идеальной клетки, необходимо принять во внимание отдельные ионные токи, протекающие через мембрану. Входящий натриевый ток (iNa) зависит от величины движущей силы для ионов натрия (Vm-ENa), а также от натриевой проводимости мембраны (gNa). Проводимость пропорциональна среднему количеству натриевых каналов, находящихся в открытом состоянии при потенциале покоя: чем больше открытых каналов, тем выше проводимость. Таким образом, натриевый ток равен:
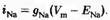
Тот же подход справедлив и для калия и хлора:
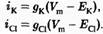
Если принять iCl, = 0 (т.е. ионы хлора находятся в равновесии), то, для сохранения неизменного значения мембранного потенциала, калиевый и натриевый токи должны быть равны по величине и противоположны по направлению:
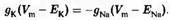
Рассмотрим это уравнение более подробно. Предположим, что gK значительно больше gNa. Тогда, для того чтобы токи были равны, движущая сила для выхода калия должна быть значительно меньше движущей силы для натриевого входа. Другими словами, мембранный потенциал должен быть ближе к ЕK, чем к ENa. Соответственно, если значение gNa велико, то мембранный потенциал будет ближе к ENa.
Преобразовав выражение, получим формулу для мембранного потенциала:
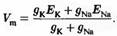
Если по той или иной причине ионы хлора не находятся в равновесии, то формула приобретает несколько более сложный вид:
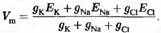
Вышеприведенные расчеты были первоначально проведены Голдманом) и, независимо от него, Ходжкином и Катцем), с той лишь разницей, что вместо равновесных потенциалов и проводимостей они оперировали ионными концентрациями снаружи ([Na] 0,. .) и внутри ([Na] i,. .) клетки, а также проницаемостью мембраны для каждого из ионов (pNa···):
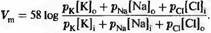
Так же как и ранее, хлорные компоненты выражения не рассматриваются, если ионы хлора находятся в равновесии. Уравнение называется "уравнением ГХК" по фамилиям авторов, или уравнением постоянного поля,
поскольку одним из допущений при выводе уравнения является равномерное распределение градиента напряжения (т.е. "поля") на всем протяжении мембраны. Уравнение ГХК полностью аналогично уравнению проводимостей, на его основании можно сделать те же предсказания: когда проницаемости для натрия и хлора малы по сравнению с калиевой проницаемостью, хлорный и натриевый компоненты уравнения становятся пренебрежимо малы, и мембранный потенциал стремится к значению равновесного потенциала для калия: Vm = 58 log ([K] 0/ [K] 1). При увеличении натриевой проницаемости мембранный потенциал сдвигается в сторону натриевого равновесного потенциала.
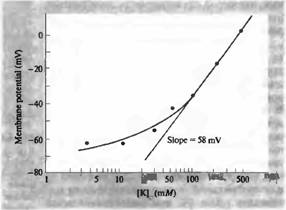
Рис.1. Зависимость мембранного потенциала от внеклеточной концентрации калия на аксоне кальмара (полулогарифмическая шкала). Прямая линия проведена под углом наклона, равным 58 мВ на десятикратное изменение внеклеточного уровня калия, в соответствии с уравнением Нернста. Благодаря наличию мембранной проницаемости для натрия, экспериментальные данные расходятся с прямой линий, особенно при низких концентрациях калия.
Из уравнения постоянного поля следует один важный принцип. Мембранный потенциал зависит от проводимости (или проницаемости) мембраны для важнейших ионов, а также от их равновесных потенциалов. В реальных клетках проводимости для калия и хлора сравнительно· высоки, поэтому потенциал покоя клетки близок к равновесным потенциалам для этих двух ионов. При увеличении натриевой проводимости, например, во время потенциала действия или возбуждающего постсинаптического потенциала, мембранный потенциал сдвигается в сторону натриевого равновесного потенциала.
|


