|
Физиологические эксперименты показывают, что активность нейронов играет определенную роль в исчезновении синапсов, влияя как на интенсивность, так и на и результат конкуренции между терминалями аксонов. Стимулирование мышцы через нерв при помощи имплантированных металлических электродов увеличивает интенсивность исчезновения синапсов. Уменьшение активности путем добавления тетродотоксина в пространство вокруг нерва для блокирования потенциалов действия приводит к ингибированию синаптической передачи и замедлению исчезновения синапсов. На мышцах, которые получают сигналы от аксонов двух различных нервов, возможно проведение интересных экспериментов по блокированию проведения импульсов избирательно в одном из нервов. В таких случаях неактивные нейроны не могут, очевидно, нормально конкурировать: аксоны блокированных нейронов иннервируют двигательных единиц меньше, чем в норме; аксоны активных нервов иннервируют волокон больше, чем обычно. Однако доминирование неблокированных нервных волокон не полное, что говорит о роли других факторов, кроме активности, в исчезновении синапсов.
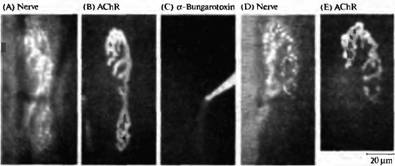
Рис. 5. Конкуренция между веточками одного мотонейрона, зависящая от активности. (А, В) Флуоресцентные микрофотографии нервно-мышечного соединения грудинососцевидной мышцы мыши, показывающие пресинаптическую терминаль (А), окрашенную при помощи 4-Di-2-Asp (который окрашивает митохондрии) и постсинаптическую мембрану (В), окрашенную при помощи родамин-связанного бунгаротоксина в низкой дозе (который окрашивает рецепторы АХ). (С) Рецепторы АХ в нижней части соединения блокированы при помощи насыщающей дозы немеченного -бунгаротоксина, что приводит к блокаде нервно-мышечной передачи селективно в этой области синапса. (D, Е) Через 31 день тот же самый синапс, что показан на А и В, был заново исследован. Терминаль аксона (D) и постсинаптические рецепторы (Е) исчезли из блокированной области. Таким образом локальная блокада передачи приводит к локальному исчезновению синапса.
Конкуренция в зависимости от активности также происходит на уровне веточек отдельного аксона двигательного нейрона. Если в небольшой области зрелого синапса произвести точечную аппликацию бунгаротоксина, то неактивный регион синапса подвергается элиминации (рис. 5). Если же блокировать весь синапс, его исчезновения не происходит. Молекулярные механизмы этой конкуренции, а также механизмы, посредством которых активность влияет на исчезновение синапсов, пока не открыты.
Сходная конкуренция В этом эксперименте среда во всех трех секциях изначально содержала ФРН. Клетки помещались в центральную секцию, а их отростки направлялись в каждую из двух боковых секций. ФРН затем удалялся из одной из боковых секций. Аксоны в центральной секции и в боковой секции, которая содержала ФРН, выживали, а аксоны в секции, из которой был удален ФРН, подвергались дегенерации (рис. 4С). Таким образом, в пределах ткани-мишени конкуренция между терминалями аксона за ограниченное количество нейротрофинов может приводить к изменениям синаптического ветвления аксонов, способствовать росту одних веточек и дегенерации других.за синаптические мишени возникает во время развития нервных путей в ЦНС. Примером может служить образование глазодоминантных колонок в зрительной коре, где аксоны из ядра ЛКТ, несущие информацию от разных глаз, первоначально значительно перекрываются в слое 4 коры, но затем формируются колонки правого и левого глаза. Таким образом, паттерн активности терминалей от обоих глаз играет решающую роль в определении результата этой конкуренции.
|


